Введение
Варрооз остается важной проблемой для пчеловодства. Поэтому изучение возможности распространения возбудителя этого заболевания — клеща варроа — различными пчелиными по-прежнему имеет большое значение.
Поведенческие реакции данного клеща ранее уже изучались отечественными и зарубежными исследователями (Непомнящих, 1980, 1981, 1983; Пилецкая 1990; Акимов и др., 1993;Calderon, Kuenen, 2001).Причем проводили исследования воздействия на поведение варроа физических и химических факторов (Гробов, 1990; Таций, 1981 и др.) либо изучали непосредственно поведенческие акты клеща и, особенно, его репродуктивное поведение, которое подробно описано разными авторами (Акимов и др., 1988, Садов, 1980; Непомнящих, 1981, 1983; Акимов и др., 1993). Практически не изученными оставались проблемы аттрактивности хозяина для клеща. Частично этот вопрос затронут в работе В. М. Та-ция (1981), однако он в первую очередь изучал реакцию клеща на градиент температур.
Возможность распознавания клещом хозяина среди другихApoidea,а также потенциальная возможность распространения варроа дикими пчелиными при контакте с медоносными пчелами изучены крайне недостаточно. В то же время поддержание культуры варроа в зимний период, когда отсуствует расплод пчел, до сих пор не представляется возможным. В связи с этим необходимо было проверить возможность развитияVarroa destructor Anolerson, Truoman,2001 на куколках диких пчелиных, зимующих в этой стадии.
Нами были проведены опыты по исследованию сравнительной аттрактивности для клеща Varroa destructor таких пчелиных, как Osmia rufa, Megachila rotundata, Bombus terrestris иApis mellifera (основного хозяина), а также изучено поведение клеща на вышеперечисленных Apoidea и репродуктивное поведение фертильных самок варроа, подсаженных на извлеченых из коконов куколок O. rufa.
Материал и методы
Материалом исследований служили клещи V. destructorи пчелы A. mellifera экспериментальной пасеки Института зоологии им. И. И. Шмальгаузена НАН Украины.
Опыты по аттрактивности клеща к различным хозяевам проводили при температуре 35°С и влажности 80%. Клещей осторожно извлекали из-под стернитов имаго пчел опытных семей и помещали в чашку Петри на растоянии 3 см от механически зафиксированных основного (A. melli-fera) и альтернативного (O.rufa, M. rotundata, B. terrestris) хозяев, расположенных под углом в 60°. Расстояние в 3 см определили опытным путем как максимальное, при котором клещ проявлял уверенный таксис к хозяину (A. mellifera). Затем фиксировали время, за которое самка клеща осуществляла выбор потенциального хозяина, закреплялась на нем. Наблюдали также за ее дальнейшим поведением на хозяине.
Результаты
Поведениеклеща. В опыте по определению аттрактивности таких видов пчел, какO. rufaиA. melliferaклещи в 42 случаях из 50 выбралиA. mellifera.Сходные результаты получены нами и при использовании в качестве альтернативного хозяина мегахил и шмелей.
Как видно из таблицы 1, шмели являются менее привлекательными для клеща варроа, чем осмии и мегахилы, однако различия не являются достоверными.
В условиях эксперимента клещи обычно вначале останавливались в характерной позе «ориентации» (Непомнящих, 1983) или «слежения» (Таций, 1981): передние ноги подняты, раздвинуты на расстояние в 1,1—1,4 мм и слегка подрагивают. Затем клещ начинает передвижение к хозяину по характерной синусоидальной траектории, близкой к описанной Р. Хайндом (1975) для телотаксиса членистоногих на примере раков-отшельников и медоносных пчел.
Клещи с удаленными передними конечностями на хозяина не реагировали.
Таким образом, можно с высокой долей вероятности предполагать о наличии механизма распознавания клещем имаго хозяина. Возможно, что он обусловлен восприятием самкой варроа одорологических, или вибрационных стимулов. Возможно также, что восприятие данным паразитом хозяина является комплексным, однако данный вопрос требует дальнейшего изучения.
Поведениехозяина. Нами проведено наблюдение за поведением V.destructor и ответными реакциями основного (A. mellifera) и альтернативного(O. rufa) хозяев во время и после закрепления на них клеща.
Подойдя к механически фиксированному хозяину и по лапке взобравшись на грудь, голову или брюшко пчелы, в 73% случаев клещ перемещался на его дорсальную сторону, в 27% — на вентральную. Однако, постепенно передвигаясь по телу пчелы, через 1 ч 82% клещей оказывались на брюшной стороне, а через 2 ч — 94%. Клещ, как правило, продолжал перемещаться, пока не оказывался под стернитами брюшка или, в ряде случаев, с дорсальной стороны груди.
 Наши наблюдения подтверждают данные В. А. Непомнящих (1983) о том, что существует рефлекс остановки клеща на опушенных покровах хозяина. Так, на значительно более опушенной, чем A. mellifera, O. rufa, общее время передвижения клеща по телу пчелы составило (10 ± 4,5) мин за 2 ч наблюдений, тогда как наA. mellifera— (18 ± 6,2) мин. При этом как медоносная пчела, так и осмия при передвижении клеща проявляли активные реакции аутгруминга.
Наши наблюдения подтверждают данные В. А. Непомнящих (1983) о том, что существует рефлекс остановки клеща на опушенных покровах хозяина. Так, на значительно более опушенной, чем A. mellifera, O. rufa, общее время передвижения клеща по телу пчелы составило (10 ± 4,5) мин за 2 ч наблюдений, тогда как наA. mellifera— (18 ± 6,2) мин. При этом как медоносная пчела, так и осмия при передвижении клеща проявляли активные реакции аутгруминга.
Таблица1.
Продолжительность выбора жертвы клещем
|
Параметры выбора
|
Apis mellifera
|
Osmia rufa
|
Megachila rotundata
|
Bombus terrestris
|
|
Количество клещей, %
|
83-92%
|
16%
|
17%
|
8%
|
|
Продолжительность выбора, с
|
44,6±14,2
|
78,87±2 6,1*
|
82,4 ±21,3**
|
243,3±35,6
|
Примечание. Коэффициент корелляции между видом жертвы и прожолжительностью выбора составляет 0,72 для Osmia rufaи 0,78 для Megachila rotundata. * Разница сApis mellifera является достоверной с р > 0,05. ** Разница с Apis mellifera является достоверной с р > 0,01.
Находясь на осмиях и несмотря на меньшую активность передвижения, клещ проявлял беспокойство, часто становился в позу ориентации с поднятой вверх первой парой конечностей, подобно описанной В. А. Непомнящих (1983), и в среднем через 24 мин покидал насекомое. Максимальное время нахождения клеща на O. rufa — 42 мин, минимальное — 9.
На пчелах, помещенных в чашку Петри без механической фиксации, для наблюдений за реакциями аутгруминга вне зависимости от положения клеща реализовывался весь комплекс движений груминга. Вначале — груминг усиков первой парой ног, затем груди и крыльев 1-й и 2-й парами, затем — вентральной и дорсальной стороны брюшка 2-й и 3-й парой ног. В зависимости от силы раздражителя цикл мог повторяться несколько раз. Наблюдалась и обратная (от брюшка к усикам) последовательность движений, однако в их структуре отличий нами не выявлено. Следует отметить, что со временем интенсивность груминга резко падала (рис. 1).
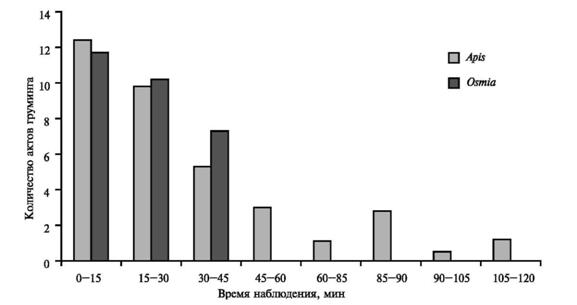
Таким образом, установлено, что интенсивность реакций груминга, выраженная количеством его циклов, практически линейно понижалась в течение первого часа, оставаясь затем на стабильно низком уровне. Возможно, это объясняется развитием охранительного торможения в ЦНС насекомого, вызванного длительным раздражением покровов клещом. Достоверных различий в интенсивности груминга между A. mellifera и O. rufaа нами не выявлено.
Развитиеклеща на куколках О.гы/а. Во второй части опыта мы проверили возможность развития клеща варроа на куколках O. rufa, отобранных на базе Института зоологии в Теремках (Киев). В конце июля нами было извлечено из коконов и помещено в пробирки при температуре 35°С 25 куколок O. rufa. Все они имели слабопигментированные покровы. В качестве контрольной группы использованы 25 куколок пчел в стадии «белоглазой куколки», предположительно находившихся на одной стадии развития с куколками осмий. Необходимо отметить, что O. rufa зимует в виде куколки с пигментированными покровами, т. е. на момент отбора их развитие продолжалось. К каждой куколке (как опытной, так и контрольной групп) подсадили самку клеща варроа, извлеченную из недавно запечатанного расплода экспериментальных семей пасеки, но еще не начавшую яйцекладку.
В течение 7 сут ежедневно мы проверяли наличие яиц клеща на куколках или стенках пробирки опытной и контрольной групп. Через 4 сут в контрольной группе 92% самок начали яйцекладку. Развитие выведшихся особей проходило нормально. В то же время в опытной группе ни одна из самок варроа яиц не отложила. Отсутствие же яйцекладки у 2 особей клеща в контрольной группе, вероятно, обусловлено заходом в ячейки перед запечатыванием самок, которые уже провели 2—3 яйцекладки и истощили возможности репродуктивной системы (Акимов и др., 1988, 1993). Вопрос о питании клеща на осмиях требует дальнейшего изучения, хотя актов питания клеща на куколках O. rufa мы не наблюдали.
Заключение
Полученные нами данные свидетельствуют о том, что клещ Varroadestructor обладает достаточно эффективным механизмом распознавания хозяина. Ввиду этого маловероятной представляется возможность его распространения и передачи одиночными пчелами при случайных контактах в природе с зараженными медоносными пчелами.
Нахождение клеща на теле пчелы в условиях опыта вызывает активные реакции аутгруминга, постепенно затихающие в течение одного часа после подсадки паразита. Реакции груминга являются схожими для различных представителей надсемейства Apoideaи отличаются значительным консерватизмом.
Доказано отсутствие яйцекладки фертильных самок клеща варроа на куколках осмий, находящихся в аналогичных с куколками A. mellifera условиях и стадии развития.
Таким образом, развитие V. destructor на одиночных пчелах не происходит не только из-за различий жизненного цикла с таковым у A. mellifera, а также вследствие определенного механизма распознавания клещом основного хозяина как в фазе имаго, так и куколки. Изучение такого механизма их распознавания представляет значительный интерес, так как использование принципа аттрактивности-репеллентности клеща к хозяину может оказаться перспективным для разработки варроацидных препаратов с механизмом действия, принципиально отличным от такового у ныне существующих.
Источник: Vestnik zoologii, 40(1): 85-88, 2006



